Beweise für DNA als Erbsubstanz
Bakterientransformationen

1928: Griffith: Transformation entdeckt
1944: O. T. Avery: chemische Natur des transformierenden Prinzip der DNA
bewiesen. Er gilt als Begründer der molekularen Genetik.
Beweis für die Transformation (Griffith):
Pneumokokken sind die Erreger der Lungenentzündung. Es existieren
verschiedene Pneumokokken-Rassen:
- S-STAMM: Wildstamm, s = smooth, haben eine Kapsel, sind pathogen (
= infektiös)
- R-STAMM: Mutanten, r = rough), haben keine Kapsel, sind nicht pathogen,
harmlos

Avery: Er hat den S-Stamm abgetötet, DNA der toten
S-Stamm-Bakterien isoliert und zu den R-Stamm-Bakterien gegeben. Diese neuen
R-Stamm-Bakterien hat er Mäusen gespritzt. Einige der Mäuse sind
gestorben.
Was ist passiert?
Die R-Stamm-Bakterien haben die DNA der S-Stamm-Bakterien aufgenommen. Durch
den Einbau der fremden DNA und deren Synthese bei der Replikation können
einige R-Zellen nun Kapseln bilden. Sie wurden zu gefährlichen S-Stamm-Bakterien
transformiert (=horizontaler Gentransfer). Diese Eigenschaft wird auch an
die Nachkommen vererbt (=vertikaler Gentransfer). Die Kapseln bestehen aus
Polysaccariden und nicht aus Nucleinsäuren. Das bedeutet, dass die
DNA die Erbsubstanz sein muss. Sie dringt in den R-Stamm ein und ändert
dort den Geno- und Phänotyp (Hülle).
Erster Beweis für DNA als Erbsubstanz!
Horizontaler Gentransfer: von einem Individuum zum Nachbarindividuum
Vertikaler Gentransfer: Gen wird an Nachkommen weitergegeben
Virusinfektion
1952: Hershey und Chase: Virusinfektion
Verwendung von Bakteriophag T2. Ein Virus, welcher das Bakterium E. Coli
befällt.
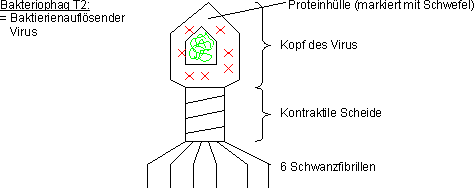
Modell eines T2-Bakteriophagen, der gerade seine DNA durch die Zellwand
in eine Bakterienzelle injiziert:

Doppelmarkierung von T2:
- Phosphormarkierung (P) der DNA
- Schwefelmarkierung (S) der Proteinhülle
Voraussetzungen: Die Proteinhülle der Viren enthält kein Phosphor
und die DNA enthält kein Schwefel. E. coli enthält kein Phosphor
in sich!
E. coli wird von T2 befallen. Die Hülle von E. coli wird aufgebrochen
und die DNA gelangt in E. coli. E. coli integriert die Phagen-DNA und synthetisiert
hunderte von neuen T2-Viren mit Proteinhüllen (solange bis E. coli
platzt). Die neusynthetisierten Phagen befallen dann neue E. coli.
Beweis:
Nur die DNA der Viren drang in E. coli ein, nicht aber die Proteinhülle.
Dies kann mit Sicherheit festgestellt werden, da sich nach dem Virenbefall
zwar Phosphor in E. coli finden liess, aber kein Schwefel, mit welchem die
Hülle markiert worden war.
Da sich in E. coli neue Phagen bildeten und nur DNA eindrang, muss also
die DNA die Erbsubstanz sein.
Chemischer Aufbau und Struktur der Nucleinsäuren
DNA und RNA
1871: F. Miescher: Hat in Basel Nucleinsäure in Lachspermien und Lachseizellen
entdeckt.
Anfangs glaubte man nicht, dass es möglich ist, dass das ganze Leben
(Prokaryonten, Eukaryonten, usw.) nur mit der DNA, also mit den vier Basen,
als Code terminiert ist. Heute weiss man, dass es tatsächlich so ist.
Die vier Basen dienen quasi wie Morsezeichen als Grundlage für die
Codierung.
Die Bausteine der Nucleinsäuren sind Nucleotide.
Bau der DNA
a) Doppelhelix
Die Doppelhelixstruktur wurde 1953 von Wilkins und R. Franklin entdeckt.
Sie haben röntgenographisch die Periodizität an kristalliner DNA
festgestellt.
› Durchmesser (20 Å) und Ganghöhe (30 Å) bestimmt!
b) Basenanalyse
Die Basenanalyse wurde 1950 erstmals von Chargaff durchgeführt. Chromatographie
und Elektrophorese nach enzymatischer Spaltung.
› A : T = G : C = 1 : 1
Basenanalyse der DNA (nach Chargaff, 1950):

c) Paarungsbedingungen

Dieser Abstand muss stimmen. Bei einer anderen Kombination der Basen könnte
der Abstand nicht eingehalten werden. Er wäre entweder zu gross oder
zu klein.
Siehe auch: DNA-Molekülkette (nachzuschlagen im Löffler oder
einem anderen Biochemiebuch)
d) Das Watson-Crick-Modell
Stoische Bedingungen für H-Brücken:
A mit T › 2 H-Brücken
C mit G › 3 H-Brücken
1953: Watson-Crick: Postulierten ein DNA-Modell: Doppelhelix

Replikation der DNA
Die Replikation erfolgt Y-förmig = semikonservative Replikation
Dank der Doppelhelix sind alle Infos doppelt (antiparallel) vorhanden. Bei
der semikonservativen Replikation bleibt die Hälfte der mütterlichen
DNA in der DNA der Tochter erhalten.
Verlauf der Y-förmigen Replikation:
- Lösen der H-Brücken zwischen den Basen. Es handelt sich um
Einfach-Ketten, für diesen Vorgang ist ein Enzym notwendig.
- Ergänzen der Basenpaare nach der Komplementaritätsregel A
= T , G = C (Enzym: DNA-Polymerasen)
- Replikation nach dem Prinzip 5‘ › 3‘
› Es entstehen zwei identische Tochterstrukturen!
Der ganze Replikationsvorgang muss sehr schnell gehen, da sonst viele
Mutationen auftreten! Bei Eukaryonten beträgt die Geschwindigkeit der
DNA-Replikation rund 50 Basen pro Sekunde (bei sehr hoher Präzision).
Bei Prokaryonten sind es 300 Basenpaare pro Sekunde.
Die Doppelhelix ist in sich verdreht. Bevor die Replikation beginnt, müssen
sich die DNA-Stränge erst noch zurückdrehen, um sich zu entwirren.
Die Replikation erfolgt immer von 5‘ nach 3‘:

Theoretisch existieren drei Replikationsarten:
- Semikonservative: ursprüngliche Doppelstruktur teilt sich in zwei
identische Tochterstrukturen
- Konservative: ursprüngliche Doppelstruktur bleibt erhalten (elterliche
Stränge), Tochterstränge entstehen neu
- Dispersive: Teile beider Stränge werden für beide Tochterstrukturen
benutzt (würde Brüche bedingen)
Beweis für die semikonservative Replikation:
1958: Mendelson/Stahl
- E. coli wir auf 15N (schweres Isotop, nicht radioaktiv) kultiviert:
es entsteht schwere DNA.
- Die erste und zweite Replikation geschehen in einem 14N-Medium.
- Ultracentrifuge: im CsCl (Cäsiumchlorid) Gradient ist die 15N-DNA
von der leichteren 14N-DNA trennbar.
Resultate für die 3 Replikationsarten:
- Semikonservative Replikation:

- Konservative Replikation:


› Wird bei jeder Replikation etwas leichter!
Experimentell konnte gezeigt werden, dass es sich bei E. coli tatsächlich
um eine semikonservative Replikation handelt.
Auch Eukaryonten replizieren semikonservativ: 1 Chromatide = 1 DNA Doppelhelix.
Geschwindigkeit der Replikation:
- E. coli: 300 Basenpaare pro sec.
- Eukaryonten: 50 Basenpaare pro sec.
Leistungen des DNA-Modells
- Längsdifferenzierung der Erbsubstanz
- Lineare Anordnung der Gene auf dem Chromosom
- Lineare Anordnung innerhalb der Gene (siehe auch Kap. 5.2: Cistron)
- Beliebig viele Kombinationen möglich
- Annahme: Ein Gen ist 2500 Basenpaare lang (enthält 2500 Basenpaare).
Alle Formeln der Natur sind mit 4 Basen codiert.
› 42500 verschiedene mögliche Kombinationen
(= 101500 versch. Kombinationen)
- Identische Replikationen
- Die Erbsubstanz repliziert sich in der Regel fehlerlos (hohe Präzision
durch semikonservative Replikation). Äusserst selten kommt es
zu einem Fehler. Dieser Fehler bedeutet eine Mutation.
Die Replikation erfolgt in der S-Phase (vor Mitose/Meiose)!
|
