Parallel verlaufende Nervenfasern
bilden im ZNS Bahnen, ausserhalb des ZNS Nerven. Jede erregbare Nervenfaser
(Axon) kann unabhängig von ihren Nachbarn Informationen in Form von
elektrischen Impulsen (AKTIONSPOTENTIALE) fortleiten.
Die erregbare Nervenfaser
Form und Dauer aller Aktionpotentiale (AP) sind meistens gleich (Dauer:
ca. 1 ms, Amplitutde ca. 120 mV). Die Intervalle zwischen den AP’s
sind verschieden lang. In ihnen ist die Information, die übertragen
werden soll, ver-schlüsselt. Eine solche Verschlüsselung wird
als Impulsfrequenzmodulation bezeichnet. So bedeutet beim Muskel eine tiefe
Frequenz eine kleine Anspannung, eine hohe eine grosse Anspannung.
Current Clamp Experimente
(beim Tintenfisch, Durchmesser des Axon = 0.5 mm)
Versuchsanordnung bei einer Membran des Axons mit variabler Stromstärke.
Die Stromstärke wird manuell verändert. Wenn Rechteckströme
kleiner Stromstärken durch die Membran fliessen, reagiert sie mit einer
kleinen lokalen Antwort. Sobald eine bestimmte Stromstärke überschritten
wird (Schwelle), entsteht ein Aktionspotential. Es gibt keine AP’s
halber Grösse. Dieses Phänomen wird als Alles-oder Nichts-Gesetz bezeichnet.
Voltage Clamp Experimente
Versuchsanordnung zwingt der Membran eine Spannung auf und der Strom, der
dazu notwendig ist, wird gemessen.
Rechteckförmige Spannungsabschnitte werden dem Ruhepotential überlagert,
die je nach Amplitude und Polarität die Membran verschieden stark hyper-
oder depolarisieren. Bei Hyper-polarisation treten Membranströme auf,
die ungefähr dem Ohmschen Gesetz folgen. Bei Depo-larisation tritt
vorerst ein Einwärtsstrom auf, dem ein Auswärtsstrom folgt. Bei
starker Depolarisa-tion der Membran wird der Einwärtsstrom kleiner
und geht bei ca. 120mV Depolarisation in einen Auswärtsstrom über.
Der Auswärtsstrom wird hingegen grösser, je stärker die Membran
depolarisiert wird.
Na+- Strom durch Tetradoxin (TTX): wird grösser
bis zu 60 mV, geht dann zurück und ändert Vorzeichen
K+-Strom durch Tetraäthylammonium (TEA):
K+ Gleichgewichtspotential ist sehr stark negativ = -80 mV, wird
die Membran depolarisiert, geht das K+-Gleichgewicht weiter
weg;
K+ -Fluss wird immer grösser
Versuche mit diesen Substanzen zeigten, dass der Membranstrom aus Na+
und K+ -Strömen besteht. Der kurzdauernde Einwärtsstrom
besteht aus Na+-Ionen, der langdauernde Auswärts-strom aus
K+-Ionen.

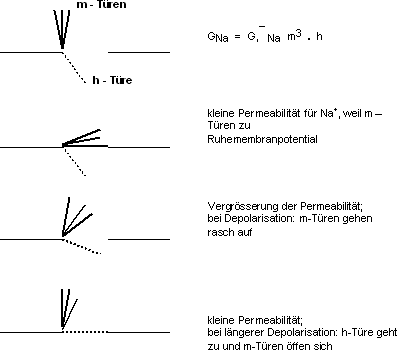
GK: Es gibt Membranproteine mit Kanälen in der Membran,
durch die nur K+-Ionen treten können. Jeder Kanal kann durch Konfigurationsänderungen
auf vier Arten verschlossen werden. Jede dieser Möglichkeiten stellen
wir uns als Türe vor. Jede „Türe“ kann also den Kanal
verschliessen. Die Wahrscheinlichkeit, dass eine „Türe“
offen ist, ist n. Daraus folgt, unter der Annahme, dass die Leitfähigkeit
GK proportional den offenen Kanälen ist:

Bei Ruhepotential sind die meisten „Türen“ geschlossen,
bei Depolarisation öffnen sie sich mit einer bestimmten Geschwindigkeit.
Türen sind “spannungsabhängig”.
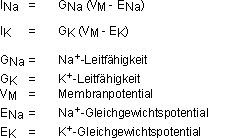
GNa: Es gibt Membranproteine mit Kanälen, durch die nur
Na+-Ionen treten können. Jeder kann durch vier „Türen“
verschlossen werden: 3 m-“Türen“ (Wahrsch.., dass offen
= m) und eine h-“Türe“ (Wahrsch., dass offen = h). Daraus
folgt:

Bei Ruhepotential sind die h-“Türen“ zu 60% offen, die
m-“Türen“ geschlossen. Bei Depolarisation öffnen sich
die m-“Türen“ rasch und die h-“Türen“
schliessen sich langsam.
Na+-Kanal
Ein Na+-Kanal kann auf vier verschiedene Arten verschlossen
werden. Jede der Türen kann den Kanal verschliessen.
› Nur wenn alle Türen offen sind, ist die Membran
permeabel.
K+-Kanal
Man hat vier gleiche Türen.
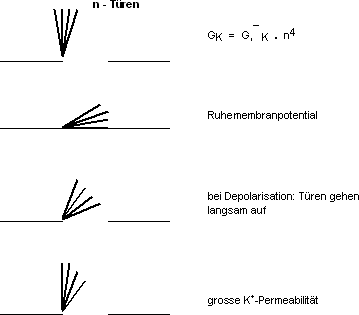
Für jeden Kanal gilt das Alles-oder Nichts-Gesetz, d.h. er ist offen
oder geschlossen. Dieses Alles- oder Nichts-Gesetz hat nichts, mit dem AP
zu tun.
Alles- oder Nichts-Gesetz des AP
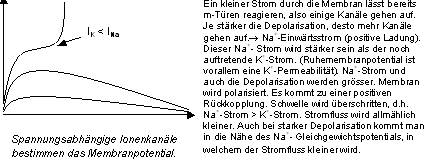
Eine Erhöhung der Reizstärke um 0.02% genügt, um aus einem
unterschwelligen (ohne AP) einen überschwelligen Reiz (mit AP) zu bekommen.
Dieses Phänomen erklärt sich mit der depolarisierenden Wirkung
des Reizes. Weil sich die m-Türen rasch öffnen, verändern
sich dabei die Ionenstärke. Sobald die Depolarisation so gross wird,
dass der Na- Einwärtsstrom grösser wird als der K-Auswärtsstrom,
tritt eine positive Rückkoppelung ein, die ein AP einleitet.
Fortleitung des Aktionspotentials
Im intakten Organismus werden AP’s vom initialen Segment zum Teledendron fortgeleitet. Es entstehen dadurch Ströme entlang des Axons. Ströme
fliessen extrazellulär zur Stelle, wo sich das AP befindet. Dadurch
werden die benachbarten Stellen depolarisiert, bis die Schwelle erreicht
und dort ein AP ausgelöst wird.
In der nichtmyelinisierten Nervenfaser muss die Fortleitung als kontinuierlicher
Vorgang ange-sehen werden. AP’s werden in dicken Fasern rascher fortgeleitet
als in dünneren, da zum einen die erregbare Membranfläche/Längeneinheit
grösser und zum anderen der Ionenwiderstand der Nervenfaser kleiner
wird (Querschnittsveränderung!) So kann Ionenstrom zweimal so gut in
Längsrichtung fliessen.
Invertebraten haben nur nicht-myelinisierte Nervenfasern, die z.T., um
grosse Fortpflanzungsge-schwindigkeiten zu erreichen (bis 30 m/s) einen
Durchmesser bis zu 0.5 mm aufweisen. Verte-braten haben neben nicht-myelinisierten
Nerverfasern auch myelinisierte, die noch grössere Fortleitungsgeschwindigkeiten
aufweisen können.
Myelinisierte Fasern
Die dicken myelinisierten Fasern haben einen Durchmesser von ca. 0.02mm.
Der Abstand zwischen zwei Schnürringen beträgt 2 mm. AP’s
entstehen nur an den Schnürringen und breiten sich saltatorisch, von
einem Schnürring zum anderen aus. Je dicker die Myelinscheide, desto
grösser der Abstand zwischen den Schnürringen, desto grösser
die Fortleitungsgeschwindigkeit. Die Myelinscheide stellt einen Ohm’schen
Isolator dar, durch den nur kapazitive Ströme fliessen.
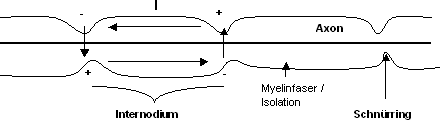
- AP wird nur an den Schnürringen generiert; deshalb “saltatorische”
Fortleitung (springend)
- Strom geht bei den Schnürringen durch ›
kapazitiver Strom, geht dann als Na+-Strom wieder rein
- Wenn die Myelinscheide dünn ist, geht viel kapazitiver Strom auf
dem Weg von Schnürring zu Schnürring verloren.D.h., es würde
ein kleiner Strom ankommen. › Bei dünnen Myelinscheidefasern
werden AP schon nach zirka 2 mm generiert, rascher also.
Refraktärperiode
Nach jeder Erregung ist eine Nervenfaser refraktär, d.h. sie ist gar
nicht oder weniger erregbar als vor der Erregung.
Absolute Refraktärperiode: Während dieser Zeit
ist die Nervenfaser unerregbar. Die absolute Refraktärperiode ist kurz
vor der Repolarisation beendet. Das Na+-System ist so stark inaktiviert,
dass der Na+- Strom nicht mehr grösser werden kann als der
K+-Strom. Die Schwelle wird so nie erreicht.
Relative Refraktärperiode: Während dieser Zeit
ist die Nervenfaser erregbar, aber die Schwelle ist erhöht. Die relative
Refraktärperiode dauert ungefär 10 ms. Während dieser Zeit
haben die h- und m-Türen ihre ursprünglichen Werte noch nicht
erreicht.
Verkleinerung von h: Inaktivierung des Na+-Systems
Vergrösserung von n: Aktivierung des K+-Systems
Dies bewirkt eine Erhöhung der Schwelle und eine Verkleinerung der
Amplitude des AP’s.
Reizzeit - Spannungskurve (Stromkurve)
Die Kurve gibt die Zeit bzw. die Stromstärke an, die benötigt
werden, um ein AP auszulösen. Spezielle Werte sind:
Rheobase: Die minimale Reizstärke eines beliebig
langen Rechteck-Reizes,
die notwendig ist, um ein AP auszulösen.
Chronaxie: Dauer eines Rechteck-Reizes doppelter Reobasen-Stärke,
der
notwendig ist, um ein AP auszulösen.
Die Reizzeit-Spannungskurve ist die Schwellenkurve für Rechteck-Reizimpulse.
Sie weist einen hyperbelähnlichen Verlauf auf mit der Gleichung:
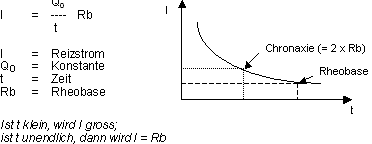
Das Summenpotential eines Nerven
Aktionspotentiale werden meist mit intrazellulären Elektroden gemessen,
wobei die Spannung zwischen Membran Innen- und Aussenseite gemessen wird.
Die Aktivität ganzer Zellgruppen kann mit extrazellulären
Elektroden gemessen werden, die nahe an die erregbaren Zellen gebracht werden.
Da extrazelluläre Ströme fliessen (s. Fortleitung
von AP) können damit Spannungen gemessen werden, die aber kleiner sind
als die transmembranären Spannungen, die ihnen zugrunde liegen. Ein
AP ist ungefähr 100 mV gross, ein SP dagegen 100 Mikrovolt, also rund
tausend mal kleiner.
Wenn zwei Reizelektroden und 2 differente Ableitelektroden am Muskelnerv
liegen, wandert eine negative Stelle (verursacht durch die individuellen
AP’s) von der Reiz- zur Ableitelektrode und weiter. Wenn sie an der
ersten und zweiten Ableitelektrode vorbei kommt, tritt jedesmal eine Spannungsschwankung
zwischen den Elektroden auf, die umgekehrt polarisiert sind. Wegen diesen
Phasen bezeichnet man das Ganze als diphasisches Summenpotential.
Es wird als Summenpotential bezeichnet, weil alle erregten Nervenfasern
dazu beitragen.
Wenn nur eine differente Ableitelektrode vorliegt (z.B. 2. Elektrode über
einen Knochen) oder der Nerv unter der einen Ableitelektrode zerstört
ist, wird ein monophasisches Summenpotential registriert.
Die Reizung eines Nerven
Wenn ein Nerv gereizt wird, ist die eine Reizelektrode negativ (Kathode),
die andere positiv (Anode). Die Nervenfasern werden an der Kathode depolarisiert
und je nach Reizstärke erregt. An der Anode werden die Fasern hingegen
hyperpolarisiert und die Schwelle ist während des Reizes dort erhöht.
Sie kann so hoch sein, dass ein fortleitendes Summenpotential blockiert
wird (Anodenblock).
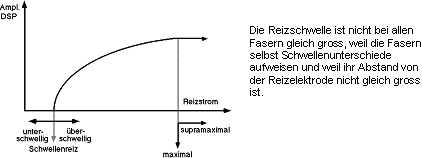
Wenn eine einzelne Nervenfaser gereizt wird, gilt das Alles- oder Nichts-Gesetz.
Bei Reizung eines Nerven ist es nicht gültig, da nicht alle Nervenfasern
die gleiche Reizschwelle aufweisen. Wenn die Reizstärke erhöht
wird, können daher 5 Reizstärken unterschieden werden, die aufgrund
der Grösse des Summenpotentials (SP) ermittelt werden:
unterschwellig, Schwellenreiz, überschwellig, maximal, supramaximal
Fortpflanzungsgeschwindigkeit des DSP
Die Zeit zwischen Reizbeginn und Beginn des DSP wird als Latenzzeit
bezeichnet. Die Latenzzeit setzt sich aus der Nutzzeit und aus der Fortpflanzungszeit
zusammen. Die Nutzzeit ist die Zeitspanne zwischen Reizbeginn und Erregungsentstehung
am Reizort. Die Fortpflanzungs-geschwindigkeit wird auf der ganzen Länge
der präparierten Nerven als konstant angenommen. Sie kann berechnet
werden, wenn die Latenzzeit von zwei verschiedenen Ableitorten bekannt ist.
Die Fortpflanzungszeit von einem Ableitungsort zum nächsten ist gleich
der Latenzzeitdifferenz.
Rasch fortleitende Nervenfaser: Rücken zu Muskeln, Muskelspindel
zu Rückenmark
Dickste Nervenfasern haben tiefste Schwelle, sind am leichtesten reizbar.
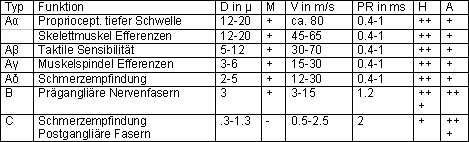
Klassifizierung der Nervenfasern
Die langsamsten Nervenfasern im ZNS des Menschen haben
eine Fortleitungsge-schwindigkeit von 1-2 m/s, die raschesten
von ungefähr 100 m/s. Die Nervenfasern werden gemäss
ihrer Fortpflanzungsgeschwindigkeit und Funktion klassifiziert.
Klassifizierung und Eigenschaften von Nervenfasern von Wirbeltieren:
D: Durchmesser, M: Myelinscheide, V: Fortleitungsgeschwindigkeit, PR: Refraktärperiode,
H: Empfindlichkeit auf Hypoxie, A: Empfindlichkeit auf Anästhesie
|
