Struktur:
- von Bindegewebshülle umgeben
- Muskelfasern durchziehen ganze Muskellänge
- Innerviert von Nerven
- Rezeptoren im Muskel: Muskelspindeln Myofibrille
Motorische Einheit:
- besteht aus Motoneuron und Muskelfasern, die mit dem Motoneuron Endplatten
bilden
- bei Erregung gilt Alles-oder-Nichts-Gesetz
Muskelfaser:
- besitzt meistens nur eine Endplatte
- Durchmesser 50-100 µm
- Muskelmembran = Sarkolem; besteht aus Einheitsmembran
(100 Å),
Polisacchariden (500 Å) und kollagenen Fasern (500 Å)
- enthält
Myofibrillen
- unter Phasenkontrast Querstreifung sichtbar
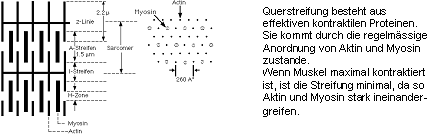
Myofibrille:
- durchzieht ganze Faser in Längsrichtung
- Durchmesser 1-2 µm
- unter Phasenkontrast Querstreifung (je nach
Dehnung Periode 2-3.5 µm)
- besteht aus Aktomyosin und ist umgeben
von transversalem Röhrensystem
und sarcoplasmatischem Retikulum
Aktomyosin:
- besteht aus den beiden kontraktilen Proteinfilamente Aktin und Myosin,
die die Querstreifung bewirken
- Aktin und Myosin greifen ineinander und
können sich gegeneinander
verschieben (Gliding filament Theory, Abb. 8.1.1)
- Entsprechend der Doppelbrechung
unterscheidet man A-Streifen, I-Streifen, H-Zone, Z-Linie
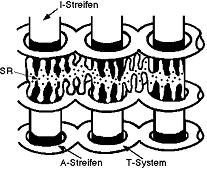
Transversales Röhrensystem (T-System):
- umgibt jede Myofibrille bei A-I-Uebergängen (Abb. 8.1.2)
- Röhren
enden im Sarkolem (offene Verbindung zwischen Lumen und extrazellulärer
Flüssigkeit)
- wichtig für die Erregung
Sarkoplasmatisches Retikulum (SR):
- bildet Verbindung zwischen zwei Niveaus vom T-System (Abb. 8.1.2)
- nimmt
13% vom Muskelvolumen ein
- auch wichtig bei Erregung des Muskel
Erregung und Kontraktion
Nach Erregung der motorischen Endplatte, finden folgende Vorgänge statt:
- EPSP löst AP aus
- AP breitet sich über Muskelfaser aus
- AP wird über T-System fortgeleitet,
wo Ca2+ –Kanäle
aktiviert werden › Ca2+ fliesst ins Sarkoplasma und wirkt auf
das SR
- SR gibt Ca2+ ab
- Ca2+ bindet sich an Aktin und ändert dessen Konfiguration
- Verbindung
der Myosin-Köpfe mit Aktin, d.h. Kontraktion geht los
- Aktivierung
der Aktomyosin-ATP-ase auf den Myosin-Köpfen
- ATP wird zu ADP abgebaut
und Myosin beugt den Hals (ergibt Kontraktion)
- ADP wird gegen ATP ausgetauscht
und das ermöglichtLösung der
Bindung zwischen Aktin und Myosin
- zeitlich unabhängig:
Reabsorption des Ca2+ durch das SR wodurch die Kontraktion
abgeschlossen wird
Es werden ca. 50 solche Zyklen/s durchlaufen. Durch das asynchrone Arbeiten
der Myosin-Moleküle kommt eine kontinuierliche Kontraktion zustande.
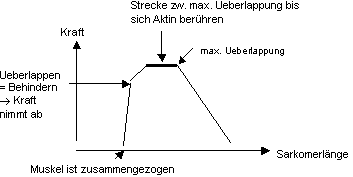
Die Kraft, die Aktomyosin entwickelt, hängt von der Sarkomerlänge
ab (Abb. 8.2), d.h. etwas vereinfacht, wie stark Aktin und Myosin überlappen
und wieviele Bindungen gebildet werden können.
Mechanik des Muskels
Bei Dehnung eines passiven Muskels steigt seine Spannung an. Die Beziehung
(Ruhe-Dehnungskurve) ist nicht linear sondern progressiv steifer, d.h. je
mehr ein Muskel gedehnt wird, desto grösser wird seine Steifheit (Abb.
8.3.1). Dieses Verhalten des Muskels kann z.T. durch folgendes mechanisches
Modell, das aus 4 Komponenten besteht, erklärt werden:
- Sehne (elastisch) = passiver Muskel
- Kontraktile Elemente (plastisch-viskös)
- Übergang Sehne-Myofibrillen
und Z-Linie (elastisch)
- Sarkolem und Bindegewebe (elastisch)
Komponente 1, 2 und 3 sind in Serie, Komponente 4 parallel zu 2 und 3 geschaltet.
Wenn sich ein Muskel aktiv kontraktiert, werden 5 Kontraktionsarten unterschieden.
Bei einer isotonischen Kontraktion bleibt die (mechanische) Spannung konstant,
und der Muskel ändert seine Länge. Die Länge der Sehne bleibt
konstant, und die oben erwähnten Komponenten 2 und 4 des Modelles verkürzen
sich.
Im Gegensatz dazu bleibt bei einer isometrischen Kontraktion die Länge
konstant, und die Spannung ändert sich. Die Sehne wird gedehnt und die
kontraktilen Elemente können sich deswegen verkürzen und entsprechend
Abb. 8.2 verändert sich dann dessen Kontraktionskraft. Unter extremen
Bedingungen, wie beim Rennen, treten sogar Situationen auf, bei denen der
Gesamtmuskel gedehnt wird, die Sarkomere sich aber simultan verkürzen.
Unterstützungs- (zuerst isometrisch, dann isotonisch) und Anschlagzuckung (zuerst isotonisch, dann isometrisch) sind zusammengesetzte Zuckungen.
Auxtonisch sind Zuckungen, bei denen sich Länge und Spannung ändern.
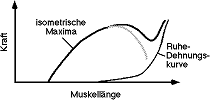
Um die isometrischen Maxima zu bekommen, wird die Spannung während
isometrischen Kontraktionen bei verschiedenen Längen aufgetragen (Abb.
8.3.2). Wegen der Dehnbarkeit der Sehne haben die Kurven auf Abb. 8.3.1 nicht
die gleiche Form.
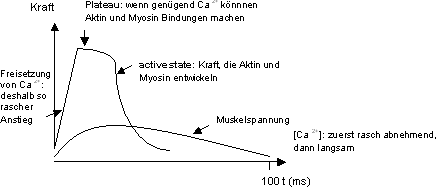
Einzelzuckung: Eine isometrische Einzelzuckung dauert je nach Muskel 30-100
ms. Die relativ grossen Beschleunigungen während der Zuckung bewirken
zusätzliche Kräfte, so dass die Spannung, die zwischen den beiden
Muskelenden gemessen wird, nicht gleich gross ist wie die Spannung zwischen
Aktin und Myosin (active state, Abb. 8.3.2). Der exponentielle Abfall des
active state verläuft parallel mit der Reabsorption des Ca2+.
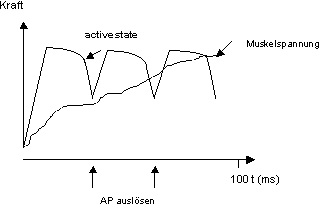
Tetanus: Wenn ein Muskel wiederholt mit genügend hoher Frequenz gereizt
wird, kann er unter isometrischen Bedingungen eine grössere Spannung
entwickeln als während einer Einzel-zuckung (Summation). Bei relativ
kleiner Frequenz kann die Spannung zwar schon grösser als bei einer
Einzelzuckung sein, es treten aber noch Spannungsschwankungen auf (unvollständiger
Tetanus, Abb. 8.3.3). Bei höherer Frequenz verschwinden die Schwankungen
und der Tetanus wird vollständig. Die Muskelspannung kann dann gleich
gross werden wie das Plateau des active state. Eine vollständiger Tetanus
kann je nach Muskel bei 20-200 Hz auftreten. Bedingung für die Tetanisierbarkeit
einer Muskels ist, dass ein AP bedeutend kürzer ist als eine Einzelkontraktion.
Die Beziehung zwischen Muskelkraft S und Verkürzungsgeschwindigkeit
v wird mit Unter-stützungszuckungen (initiale Länge = konstant)
untersucht, wobei die initiale Geschwindigkeit gemessen wird (Abb. 8.3.4).
Wenn diese Beziehung für verschiedene Ausgangslängen des Muskels
gemessen wird, so liegt die Kurve für Ruhelänge höher als
die Kurven für kleinere oder grössere Längen. Aus der Kraft-Geschwindigkeitsbeziehung
kann die Leistung (L = S x v) berechnet werden. Die Leistungskurve (Abb.
8.3.4) durchläuft ein Maximum und liegt am höchsten um Ruhelänge.
Wir können daraus schliessen, dass die Leistung eines Muskels am grössten
ist, wenn extreme Belastungen, Längen und Geschwindigkeiten vermieden
werden.
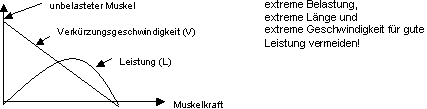
Ermüdung
In einem ermüdeten Muskel ist die Kontraktionsamplitude kleiner und
die Erschlaffungsdauer grösser als normal. Folgende Faktoren spielen
dabei eine Rolle:
- pH-Änderung wegen Milchsäureansammlung
- ATP-Mangel (Brücken
zwischen Aktin und Myosin können nicht
mehr gelöst werden). Erschlaffung des Muskels wird länger und
Stärke der Kontraktion nimmt ab.
Aktomyosin wird nicht mehr mit ATP ausgetauscht › Muskel
wird schwächer,
steifer › alles wird langsame
Kontraktur, Starre
Darunter versteht man länger andauernde Kontraktionen, die nicht
durch AP ausgelöst werden. Kontrakturen sind reversibel, Starren irreversibel.
(Muskel ist geschädigt.)
Kontrakturen:
- Depolarisations-Kontraktur (K+-Ionen, Ach) = Muskelmembran wird ohne
AP depolarisiert
- chem. Kontrakturen (ATP-Mangel, O2-Mangel)
Starren:
- Totenstarre: kein ATP mehr vorhanden! Bindungen zwischen Aktin und
Myosin sind gemacht › Muskel bleibt steiff
- Wärmestarre: Aktin
und Myosin verbinden sich durch Wärme
- Säurestarre: durch Senken
des ph-Wertes
Energetik
Der Ruheumsatz von Muskeln beträgt 1-2 Watt/kg. Sie produzieren 60%
der Körperwärme.
Für die Muskelarbeit wird ATP verwendet, das auf folgende Weise beschafft
werden kann:
rasche Beschaffung v.a. für Spitzenleistungen:
- aus Vorrat (5 mM ATP/kg Muskel reicht nur für wenige Zuckungen)
- aus
Kreatininphosphat: bei Abbau davon entsteht ATP. Das passiert während
der Arbeit. Im Ruhezustand brauchen wir ATP, um diesen Vorrat wieder
aufzubauen.
- anaerober Abbau von Zucker zu Milchsäure (Glykolyse)
Langsame Beschaffung für Dauerleistungen:
- aerober Abbau von Zucker zu ATP und CO2 (oxidative Phosphorylierung,
Wirkungsgrad 55%).
Der Wirkungsgrad bei der Umwandlung von ATP in ADP und mechanische Energie
beträgt 40%. D. h. 60% gehen in Wärme verloren. Der Gesamtwirkungsgrad
von Glucose zu mechanischer Arbeit beträgt also 20-25%.
Die vom Muskel produzierte Wärme wird unterteilt in:
Initiale Wärme (vor und während Kontraktion)
- Aktivierungswärme (grösster Teil der initialen Wärme
wird während isometrischer Einzelzuckung freigesetzt, wird v.a.
verursacht durch Freigabe von Ca 2+ und Spaltung von ATP)- Erhaltungswärme
(=Aktivierungswärme
während isometrischem Tetanus)
- Verkürzungswärme (Fenn-Effekt,
proportional der Verkürzung)
- Erschlaffungswärme (proportional
der Muskelspannung)
Erholungswärme
- braucht O2 , wird anaerob gemacht, d.h. mann muss mehr atmen
Einsatz der Skelettmuskulatur
Auf die meisten Gelenke wirken mehrere Muskeln. (Sie lassen sich in Agonisten,
die in Richtung der Bewegung, und Antagonisten, die entgegen der Bewegung
wirken, einteilen.) Etliche Muskeln wirken über mehr als ein Gelenk.
Wenn ein Muskel bei einer Bewegung aktiviert wird, hängt das Drehmoment,
das dadurch über dem Gelenk erzeugt wird, vom Hebelarm ab, der wiederum
von den Insertionsstellen des Muskels und vom Winkel des Gelenkes abhängig
ist. Wenn der Zusammenhang zwischen maximaler Kraft und Muskellänge
in vivo studiert wird, muss das Drehmoment durch den Hebelarm, der vom
Winkel abhängig ist, dividiert werden. Bei der willentlichen Durchführung
von Bewegungen werden alle diese Faktoren vom ZNS berücksichtigt.
Die Muskeln sind verschieden aufgebaut je nach den Aufgaben, für
die sie vom Körper eingesetzt werden. Folgende Unterschiede zwischen
Muskeln lassen sich mit ihren funktionellen Unterschieden in Zusammenhang
bringen:
- Anordnung der Fasern: Die Fasern sind parallel zur Zugrichtung, wenn
grosse Verkürzungen bei kleiner Last, hingegen schräg dazu,
wenn kleine Verkürzungen bei grosser Last notwendig sind.
- Dauer einer
Einzelzuckung:
Augenmuskel: 10 ms, Gastrocnemius: 40 ms, Soleus: 100 ms
- Grösse der
motorischen Einheiten (Fasern/mot. Einheit):
Augenmuskel: 10, Tibialis: 500, Soleus: 1500
- Durchmesser der Fasern:
Augenmuskel 20 µm, Tibialis 60 µm
Die meisten Muskeln sind aus drei Typen von motorischen Einheiten zusammengesetzt:
FF (fast twitch fatiguable), FR (fast twitch, fatigue resistent), S (slow
twitch). Die funktionell wichtigsten Eigenschaften der drei Typen sind
auf der folgenden Tabelle ersichtlich.

Die Kraft für eines Muskels wird vom ZNS mit zwei Mechanismen reguliert:
- Rekrutierung von motorischen Einheiten: Es gilt dabei meistens das Grössenprinzip:
für feine Bewegungen werden kleine langsame S Einheiten,
für grobe kraftvolle Bewegungen zusätzlich grössere und
raschere FR und FF Einheiten eingesetzt. Die Kraftzunahme pro motorische
Einheit bleibt auf diese Weise ungefähr konstant.
- Frequenzmodulation:
Je grösser die Muskelkraft,
desto höher
wird die Ent ladungsfrequenz der motorischen Einheiten und dadurch auch
ihre mittlere Kraftentwicklung.
Leistung des Muskels
Zu Beginn einer Dauerleistung wird eine Sauerstoffschuld eingegangen (v.a.
durch Abbau von Kreatininphosphat). Der Organismus stellt sich dann auf oxidative
Phosphorilisierung um. Im Vergleich zum „Ruhezustand“ ist während
einer Dauerleistung:
- der ATP-Abbau 100 -1000 mal erhöht.
- die Muskeldurchblutung 20 mal
erhöht.
- das Herzminutenvolumen und die Herzfrequenz 2 -3 mal erhöht.
Die maximale Dauerleistung beträgt ca. 400 Watt (0.5 PS). Sie wird
durch die oxidative Phosphorilisierung oder den Kreislauf bestimmt.
Der anaerobe Glukoseabbau (Glykolyse) während einer Spitzenleistung ist
2 -3 mal rascher als die oxidative Phosphorilisierung. Beim Start eines Sprinters
können bis zu 3 kWatt (4 PS) geleistet werden.
Überblick über die funktionelle und strukturelle Organisation der Motorik
Die Motorik des Menschen umfasst sowohl geplantes als auch instinktives Verhalten und bedeutet Interaktion mit der Umwelt. Der Mensch hat die Fähigkeit, seine Handlungen zu planen: seine Motorik ist zukunftsorientiert.
Die wichtigsten motorischen Strukturen Kortexareale:
- Primär-motorischer Kortex (Area 4) – Somatotopie
- Supplementär-motorischer Kortex (Area 6 mesial)
- Prämotorischer Kortex (Area 6 lateral)
- Frontales Augenfeld (Area 8)
- Motorisches Sprachzentrum von Brocca (Area 44 – unilateral)
- Somatosensorischer Kortex (Areae 3,1,2)
- Parietaler Assoziationskortex (Areae 5 und 7)
Absteigende Bahnen:
- Pyramidenbahn
- Extrapyramidale Bahnen
- Vegetative Bahnen
Interne Schleifen über das Zerebellum und die Basalganglien
Stehen und Gehen
Der Muskeltonus ist eine biomechanische Notwendigkeit für das aufrechte Stehen und die Grundlage der Halte- und Stützmotorik
Tonus: anhaltende Aktivität der Muskulatur, welche die Versteifung respektive Dehnbarkeit der Muskeln bestimmt
Der aufrechte Stand ist ein dynamisches Geschehen, dem eine präzise, automatische Regulation und eine Zielvorgabe zugrunde liegen
Die Stützmotorik gewährleistet die für das aufrechte Stehen kritische Körperstabilität
Die Lokomotionsrhythmen sind im Rückenmark organisiert; sie werden durch ein longitudinales neuronales Netzwerk im Hirnstamm gesteuert
Die natürliche, zielgerichtete Lokomotion impliziert auch Kleinhirn, Basalganglien und sensomotorische Kortexareale
Reflexe und propriospinaler Apparat des Rückenmarks Das Netzwerk der Interneurone im Rückenmark bildet die strukturelle Grundlage für seine Eigenfunktion; die Klassifikation der Interneurone erfolgt nach ihrer Verschaltung und ihren Aufgaben
Die Motoneurone bilden die gemeinsame Endstrecke in der Befehlsübertragung
Querschnittslähmung
Die Eigenfunktion des Rückenmarkes manifestiert sich in patho-logischer Weise bei Querschnittsverletzungen des Rückenmarkes. Nach akuter Verletzung sind kaudal von der Läsion die Körperteile völlig gelähmt und schlaff; es können auch keine somatischen und vegetativen Reflexe ausgelöst werden. Nach dieser sogenannten spinalen Schockphase, die mehrere Wochen andauert, erholen sich allmählich die Reflexe und der Muskeltonus wieder, es bleibt aber das Unvermögen der willentlichen Kontrolle der Bewegungen. Die Langzeitveränderungen können über Monate anhalten und führen zu einer ausgeprägten Erregbarkeitssteigerung, wobei auch die reflektorische Kontrolle der Sphinkteren und der Gewebedurchblutung gestört sein können.
Wer querschnittsgelähmt ist, kann sich nicht aus eigener Kraft bewegen. Aus diesem Grund sind Patienten auf Hilfsmittel angewiesen, die ihnen ihre Mobilität zurückgeben. Dazu zählt an erster Stelle der Rollstuhl, mit denen sich jemand mit einer Querschnittslähmung relativ frei bewegen kann, auch wenn er seine Beine nicht spürt. Die Umgebung muss dafür jedoch weitestgehend barrierefrei sein, denn zu enge Wege, Treppen oder unwegsames Gelände sind nicht für Rollstühle geeignet. Erleidet jemand eine Querschnittslähmung ist ein Umzug in den meisten Fällen nicht zu vermeiden. Lebt man in einem Eigenheim, muss dieses angepasst werden, indem beispielsweise ein Fahrstuhl oder Treppenlift eingebaut wird. Treppenlifte gibt es in verschiedenen Ausführungen, auch extra für den Gebrauch mit einem Rollstuhl. Schwellen müssen geebnet werden und es wird ausreichend Platz benötigt, denn zu enge Kurven stellen ein Hindernis für Rollstuhlfahrer dar. Es gibt mittlerweile die Möglichkeit Autos so umzubauen, dass sie trotz Querschnittslähmung sicher gefahren werden können, diese Umbaumaßnahmen sind jedoch ziemlich kostspielig. Mit den öffentlichen Verkehrsmitteln können Rollstuhlfahrer ebenfalls von A nach B gelangen. Busse sind in der Regel mit einer ausfahrbaren Rampe ausgestattet und an Hochbahnsteigen ist auch der Einstieg in Stadtbahnen unproblematisch. An den meisten U-Bahn-Stationen gibt es Fahrstühle, die für Rollstuhlfahrer oder auch für Personen mit Kinderwagen eingerichtet wurden. Es gibt viele Möglichkeiten sich trotz Rollstuhl nicht einschränken zu lassen. In der Therapie wird dabei geholfen und der sichere Umgang mit Hilfsmitteln gelehrt.
Reflexe sind automatische motorische Reaktionen auf äussere Störimpulse
Zum Reflexbegriff gilt die Regel, dass bei zunehmender Stärke des äusseren Reizes die ausgelöste Reflexantwort über dem Schwellenwert stetig zunimmt bis der Maximalwert (Sättigung) erreicht ist. Der Reflex ist willentlich nicht unterdrückbar.
Der monosynaptische Dehnungsreflex dient der Lagestabilisierung
Gleichzeitig nimmt die Spannung von Antagonisten des gedehnten Muskels ab; dies erfolgt über eine disynaptische Hemmung der Motoneurone über hemmende Ia-Interneurone.
Funktion des monosynaptischen Dehnungsreflexes: Konstanthaltung der Muskellänge (Längenservo).
Die Empfindlichkeit der Muskelspindel wird durch die alpha-Motoneurone von zentral gesteuert
Funktion des γ-Antriebs: Der Sensor kann unabhängig von der jeweiligen Muskellänge stets auf äußere Störungen reagieren.
Die Auslösung des monosynaptischen Eigenreflexes mit dem Reflexhammer dient der Exploration der Erregbarkeit des Reflexbogens; zur quantitativen Prüfung dient die elektrische Reizung des peripheren Nerven (Messung des H-Reflexes)
Klinische Reflexprüfung
Den Arzt interessieren allfällige Seitenunterschiede in der Intensität der Reflexantwort. Bei einer Halbseitenlähmung mit pathologischer Erhöhung des Muskeltonus (Spastizität) wird man z.B. auf der kranken Seite einen viel „lebhafteren“ Reflex auslösen können. Das Fehlen der Reflexe kann diagnostisch auf eine Unterbrechung im afferenten oder efferenten Schenkel hinweisen. Allerdings sind auch beim gesunden Menschen die Sehnenreflexe häufig nur schwach und schwer auslösbar. Der Hoffmann-Reflex
Da die Ia-Spindelafferenzen eine etwas tiefere Schwelle haben als die Fasern der alpha-Motoneurone, gelingt es bei geringer Reizstärke die Ia-Afferenzen selektiv zu erregen. Die elektrische Reizstärke kann fein dosiert und konstant gehalten werden. Die Grösse der Reflexantwort kann somit anhand des elektromyographisch registrierten Summen-potentials genau gemessen werden.
Die erzeugten Aktionspotentiale in den Motoaxonen werden bei der künstlichen Reizung sowohl in die Peripherie als auch nach zentral geleitet. Im Motoneuron kommt es dadurch zu einer „Kollision“ der zentralwärts geleiteten Impulse und der reflektorisch erzeugten Impulse, die sich gegenseitig auslöschen. Dadurch erlärt sich das Phänomen, dass mit zunehmender überschwelliger Erregung motorischer Axone im gemischten Nerven die Amplitude des H-Reflexes wieder abnimmt. Polysynaptische Reflexe (Fremdreflexe) sind über spinale Interneuronenketten mit den motorischen Einheiten verknüpft; funktionell gesehen sind sie Bausteine protektiver Reaktionen
Potentiell schmerzhafte und unerwartete Reize bewirken ebenfalls einen Fremdreiz, allerdings mit sukzessiver Abnahme der Reflexantwort bei regelmässiger Reizwiederholung. Diese für die Fremdreflexe typische Phänomen nennt man Habituation.
Variabilität polysynaptischer Reflexe. Die Fremdreflexe sind in der Latenzzeit, Dauer, Amplitude und Ausbreitung der Antwort ziemlich variabel. Dies ist der polysynaptischen Uebertragung zuzuschreiben, denn mit jeder zusätzlichen Synapse im Reflexbogen steigt die Variabilität und die Unsicherheit in der Uebertragung.
Sensomotorische spinale Integration und Hemm-Mechanismen
Die motorische Regulation erfolgt über ein kooperatives Zusammenspiel der verschiedenen Sinnesrezeptoren. Bewusste Wahrnehmungen, Körperschema und automatische Korrekturen.
Spinale Hemmechanismen unterstehen als Regelkreise einer zentralen Kontrolle: Regulation der Spannung durch die Golgi-Sehnenorgane, Motorservo und Renshaw-Hemmung
Der Reflexweg der Golgi-Sehnenorgane ist disynaptisch und schliesst ein Ib-Interneuron ein, das hemmend auf die Motoneurone zurückwirkt. Ueber diesen Regelkreis, kann z.B. bei Ermüdung des Muskels und drohendem Spannungsabfall, die Kraft durch Abnahme der Golgi-Hemmung konstant gehalten werden.
Durch bahnende oder hemmende Effekte der absteigenden Fasern können je nach Bedarf durch das Programm Schaltkreise geöffnet oder geschlossen werden („gating“-Phänomen). Ferner können durch den Mechanismus der präsynaptischen Hemmung Effekte von primären Afferenzen unterdrückt werden.
Ein Verlust der somatischen Sensibilität hat katastrophale Folgen für motorische Leistungen
Zielbewegungen des Armes und Greifen
Zielgerichtetes Greifen ist ein visomotorischer Akt mit kognitiver Komponente
Eine gezielte Handbewegung erfolgt mit variablen Bewegungsspuren, wobei die Zielgenauigkeit negativ mit der Geschwindigkeit korreliert
‚Prinzip der motorischen Aequivalenz‘: Zum Beispiel verändert sich die Individualität der Schriftzüge nur unwesentlich, egal ob wir nur mit Fingerbewegungen, der ganzen Hand oder sogar mit dem ganzen Arm schreiben.
Die hohe Sensibilität der Hand ist für die Greiffunktion unentbehrlich
Unbewusste Griffkorrekturen bei plötzlichen Aenderungen der Last oder der Haftung zwischen der Hand und der Objektoberfläche
Die Bewegungsrichtung einer Armbewegung wird durch eine Population von Neuronen des motorischen Kortex kodiert
Bei gezielten Armbewegungen verändert sich die Aktivität einer grösseren Population von Neuronen im Armareal der motorischen Hirnrinde; diese beginnt etwa 100-200 ms vor Bewegungsbeginn und klingt während der Bewegung wieder ab.
Die Resultante aller Neuronen-Einzelvektoren ergibt den Populations-vektor.
Die Hand ist in den sensomotorischen Kortexarealen sehr stark repräsentiert und die Kontrolle erfolgt über die Pyramidenbahn
Das koritkomotoneuronale System. Ein Teil der Pyramidenbahnfasern aus dem Areal MI ist mit den Motoneuronen direkt, d.h. mono-synaptisch verbunden.
Läsionsfolgen für die Greiffunktion. Bei experimenteller Durch-trennung der Pyramidenbahn des Affen manifestiert sich die motorische Störung vorwiegend am Verlust der Handgeschicklichkeit – Verlust des Präzisionsgriffes.
|
