Die Kapillarwände trennen die
interstitielle Flüssigkeit vom Plasma und bestehen aus ein-schichtigen
Endothelzellen mit Abständen. Die Kapillarwände lassen alle Plasmateile
mit Ausnahme der Proteine durch. Im Plasma hat es viele Proteine, im interstitiellen
Raum keine.
Kapillarwände
Filtration: Fluss durch eine Membran aufgrund eines unterschiedlichen
hydrostati-
schen Druckes (z.B. Blutdruck, der durch das Herz aufgebaut wird). Falls
die Membran nicht für alle Substanzen permeabel ist spricht man von
Ultrafiltration.
Diffusion: Fluss von höherer zu niedrigerer Konzentration
aufgrund der Brownschen Molekularbewegung › ungerichtete
Bewegung, z.B. Molekül in der Luft macht ständig Bewegung dadurch
entsteht ein Transport und somit ein Ausgleich der Konzentrationen ›
geht zum Gleichgewicht;
Ist die Membran für mehr als ein Ion permeabel, haben wir eine Diffusion
(bei Nettofluss).
Diffusion ungeladener Moleküle
a) Gase: Diffusion von Gasen wird bestimmt durch den Unterschied
zwischen Partialdruck (Mass für Konzentration) und Diffusionskoeffizienten
(Mass für Beweglichkeit)
z.B. O2 , CO 2 :
O2 › Lunge › Blutkörperchen
› Kapillare › aus dem Blut
in den interstitiellen Raum › in die Zelle›
von Mitochondrien gebraucht › CO 2 geht den gleichen
Weg zurück.
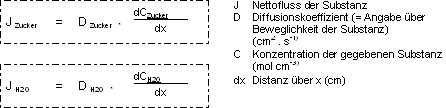
b) in H2O gelöste Teilchen: Der Fluss (J) entspricht
der Anzahl Mole pro Sekunde, welche durch eine Einheit Oberfläche wandern.
Z.B. konzentrierte Zuckerlösung in Kontakt mit reinem Wasser:
Gleichgewicht: keine Energie, Systemparameter konstant
Ungleichgewicht: Energieverbrauch, Systemparameter variabel
steady state: Energieverbrauch, Systemparameter konstant
 Der
Nettofluss wird vom Ort der grösseren Konzentration zum Ort mit der
geringeren Konzentra-tion sich bewegen, das heisst, dass er proportional
zum Konzentrationsgradienten der Substanz ist. H2O diffundiert in entgegengesetzte
Richtung. Der
Nettofluss wird vom Ort der grösseren Konzentration zum Ort mit der
geringeren Konzentra-tion sich bewegen, das heisst, dass er proportional
zum Konzentrationsgradienten der Substanz ist. H2O diffundiert in entgegengesetzte
Richtung.

Die Diffusionszeit verhält sich proportional zur Distanz im Quadrat
Þ Diffusion ist nur über kleine Distanzen effizient. Diffusion
als nicht geeigneter Substanztransporter bezüglich des ganzen Körpers.
Im Gleichgewicht, d.h. wenn die Konzentrationen gleich sind, sind die Flüsse
in beide Richtungen die gleichen, d.h. es gibt keine Nettoflüsse mehr.
Diffusionsgesetz von Fick:
In Flüssigkeiten wie dem Extrazellulärraum und dem Zytosol gleichen
sich Konzentrations-unterschiede von gelösten Stoffen durch Diffusion
aus (1. Fick Diffusionsgesetz). Diffusion durch Membran wird mit Permeabilität
beschrieben.
Der Nettofluss, das heisst J1›2 - J2›1 wird durch das Fick’sche
Diffusionsgesetz ausgedrückt (unidirektioneller Fluss):

Die Triebkraft des Flusses ist der Konzentrationsunterschied. Wenn eine
Membran für alles permeabel ist, so ist nach einer gewissen Zeit die
Konzentration überall gleich und es tritt keine Volumenänderung
ein.
(Frei durch die Lipidmembran diffundieren: H2O, gelöste Gase wie O2
und CO2, lipidlösliche Stoffe, kleine polare Moleküle wie Ethanol,
Harnstoff
Lipidschicht praktisch impermeabel für: geladene Moleküle)
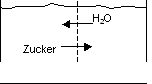
c) Diffusion von Wasser und das Phänomen der Osmose
Osmose ist Diffusion des Wassers. Antrieb ist der Konzentrationsgradient
für das Wasser selbst. (cH2O = Kehrwert der Osmolalität) Die Osmose
verursacht einen osmotischen Druck.
Wenn eine Membran für einen in Wasser gelösten Stoff undurchlässig
ist, aber durchlässig für Wasser ist, so geht das Wasser durch
(Diffusion), um den Konzentrationsunterschied von x auf der einen Membranseite
auszugleichen. Es erfolgt eine Volumenänderung.

› Bei 1 osmolaler Lösung pro Liter beträgt
der korrespondierende osmostische Druck
25,4 Atm. bei 37° C.
D.h. im Versuch steigt bei einer 1 molaren Zuckerlösung die Höhe
auf 254 m an. 1 m entspricht 25,4 Atm.
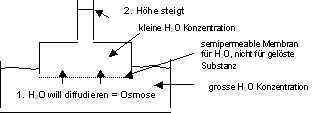
Gleichgewicht, wenn hydrostatischer Druck = osmotischer Druck
Der Körper hat überall die gleiche Osmolalität, d.h. 0,3
osmol / kg Wasser. Man nennt das isoosmotisch.
› als 0,3 osmol › hyperosmotisch
< als 0,3 osmol › hypoosmotisch
Weisen alle Kompartimente des Körpers diesselbe Osmolalität auf,
dann ist die Konzentration von Wasser konstant.
Diffusion von geladenen Molekülen (Ionenfluss)
Diffusion von geladenen Teilchen wird nicht nur durch den Konzentrationsgradienten
sondern auch durch die elektr. Spannung angetrieben. Der Fluss bewirkt eine
Ladungsänderung. Dies bewirkt wiederum ein elektrisches Feld, welches
als Spannung dem Diffusionsfluss entgegen-wirkt.

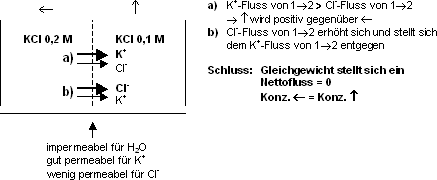
Membran, welche nur für ein Ion permeabel ist
Gegebene Situation: Membran ist nur für K+ permeabel,
[KCL] links = 0.1 mol, [KCL] rechts = 0.01 mol.
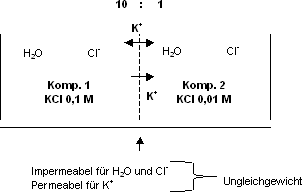
Sobald einige wenige, nicht messbare K+ - Ionen die Membrane
passiert haben, baut sich ein elektrischer Gradient auf. Die Seite 1 wird
in Bezug zur Seite 2 negativ. Wollen noch mehr K+ -Ionen diffundieren,
werden diese abgestossen durch die bereits positive Ladung, d.h. ein Teil
der K+ -Ionen bleiben im Komp. 1 › Gleichgewicht entsteht,
d.h. Nettofluss ist null. Ein Gleichgewicht zwischen chemischer und elektrischer
Kraft stellt sich sofort ein. Man spricht von einem Gleichgewichtspotential:
Nernstsche Gleichung:
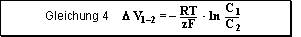
R Allg. Gaskonstante (8,31 J/Kmol)
T absolute Temperatur (273.15 K = 0°)
F Farradaykonstante (96'000 C pro mol)
z Wertigkeit des Ions (bei Cl- z.B. -1)
C1 Ionenkonzentration extrazellulär
C2 Ionenkonzentration intrazellulär
So ergibt sich z.B., dass ein einwertiges Ion mit einem Konzentrationsverhältnis
über der Membran von 10 : 1 eine Membranspannung von 60 mV erzeugt.
Membran, welche nur für mehrere Ionen permeabel ist
Die Membranspannung in Gegenwart mehrerer Ionensorten wird durch die Goldmann-
bzw. constant-field-Gleichung beschrieben (d.h. kann nur angewendet werden,
wenn elektr. Feld konstant ist). Sie gibt uns Auskunft über das Diffusionspotential
(nicht das Gleichgewichts-potential !!) zu jedem Zeitpunkt der Diffusion.
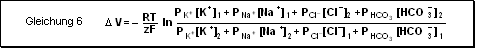
Membran, welche für mehrere Ionen permeabel, für ein Ion aber
nicht permeabel ist = Donnan-Gleichgewicht
In diesem Fall verunmöglicht das nicht-permeable Ion den Konzentrationsausgleich
zwischen den permeablen Ionen. Wir erhalten eine Gleichgewichtssituation
mit einer Potentialdifferenz zwischen den Kompartimenten.
Im Gleichgewicht müssen zwei Bedingungen erfüllt sein:
- Elektroneutralität
- Verteilung nach dem Gleichgewichtspotential
Gegebene Situation:
Kompartiment 1 : NaCl, Kompartiment 2 : NaCl + Proteine; die
Membran ist für Proteine und Wasser undurchlässig, für Na+
und Cl- jedoch durchlässig
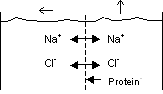
Proteine liegen im Blut bei physiologischem pH vorwiegend als Anionen vor.
Dadurch, dass bei der Ultrafiltration die Proteinmoleküle zurückgehalten
werden, ergibt sich eine Ungleichverteilung aller beteiligten Ionensorten
diesseits und jenseits der Filtermembran.
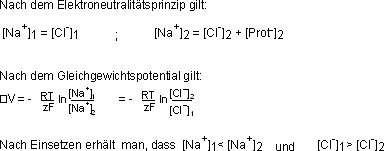
Setzt man die Konzentrationswerte von Na+ und Cl- der interstitiellen
Flüssigkeit bzw. des Plasmas in der Gleichtgewichtspotential-Gleichung
ein, so erhält man mit der Nernst-Gleichung eine Potentialdifferenz
von DV = 0.9 mV (d.h. die interstitielle Flüssigkeit ist positiv gegenüber
dem Plasma).
Die Folge dieser ungleichen Verteilung der Ionen zwischen den beiden Kompartimenten
ist ein ungleiches Teilchenverhältnis. Die Seite mit den Proteinen
hat eine grössere Osmolalität als die ohne.› Konsequenz falls
die Membran für Wasser durchlässig ist: OSMOSE. Das Plasma enthält
1,5 mosm mehr als die interstitielle Flüssigkeit, was einem osmotischen
Druck von 25 mmHg entspricht. Der resultierende Wasserfluss, von der interstitiellen
Flüssigkeit zum Plasma, wird durch den hydrostatischen Druck von 25
mmHg (durch das Herzpumpen erzeugt) in den Kapillaren ausgeglichen.
Zellmembran
Die Zellmembran ist eine sehr selektive Barriere. Die Phospolipide der
Zellmembran lassen lipophile Substanzen diffundieren, während sie für
hydrophile Substanzen eine Barriere darstellen. Für diesen Fall gibt
es die integralen Membranproteine, die als Kanäle mit einem maximalen
Durchmesser von 7 Angström dienen.
Permeabilitätsmessung
- mit radioaktiven Isotopen
- mit Strom- und Spannungsmessung
- mit Osmose, aufgrund des Wasserflusses
- Isoosmotische Lösung mit einem permeablen Solut:
Die Konzentration dieser Substanz ist aussen höher als innerhalb
der Zelle. Somit gibt es eine Nettodiffusion von aussen nach innen
mit dem Resultat, dass innen die H2O-Konzentration abnimmt und aussen
steigt. Die Zellmembran ist für H2O permeabel, es gibt einen
sekundären Wasserfluss mit der Konsequenz, dass das Zellvolumen
steigt und die Zellen platzen.
- Isoosmotische Lösung mit einem impermeabelen Solut:
Nichts passiert, keine Zytolyse oder Hämolyse, man nennt diese
Lösung auch isotonisch. Bei isotonischen Lösungen erfolgt
kein primärer oder sekundärer Wasserfluss.
isotonische Lösung = isoosmotische Lösung bei welcher das gelöste
Solut impermeabel ist
= gleicher osmotischer Druck
isoosmotische Lösung = Lösung mit gleicher Anzahl osmotisch aktiver
Moleküle auf beiden Seiten der Membran
hypertonisch = Lösung mit höherem osmotischen Druck als in der
Zelle
= hyperosmotische Lösung mit nicht permeablem Stoff › primärer
Wasserfluss › Zellen schrumpfen
hypotonisch = hypoosmotische Lösung mit impermeablem Solut › primärer
Wasserfluss von aussen in die Zelle hinein › Zellen
platzen
› Flüssigkeitsverluste beim Menschen darf man
nicht durch reines (=destilliertes) Wasser ausgleichen. Man verwendet z.B.
eine physiologische Kochsalzlösung (0,95 g Na Cl in 100 g Wasser).
Permeabilität der Zellwand | Für nicht geladene Substanzen (Passiver
Transport)
- Fettlösliche Stoffe durchqueren die Zellmembran durch einfache
Diffusion nach der Gleichung von Fick: J netx = Px × D Cx ×
A
- Wasserlösliche Stoffe kleiner als 7 A° und H2O diffundieren
durch Membrankanäle durch einfache Diffusion in Abhängigkeit
der Gleichung von Fick.
- Wasserlösliche Stoffe grösser als 7 A° (Glukose, Aminosäuren)
gelangen in die Zellen durch erleicherte Diffusion mit Hilfe von Trägermolekülen.
Im Vergleich zur einfachen Diffusion erlaubt die erleichterte Diffusion
einen schnelleren Transport dank Transportproteinen. Sie ist sättigbar
und verläuft nicht nach der Gleichung von Fick. Bis Sättigung
eintritt, vergeht Zeit!
Merkmale:
- Sättigung
- Selektivität
- Blockierbarkeit der Kanäle
- Die treibende Kraft ist ein Konzentrationsgradient ohne Hilfe von Energie
› passiver Transport
Für geladene Substanzen (hydrophil) (Passiver Transport)
Geladene Stoffe diffundieren durch die Zellmembran nach der Gleichung 3:
Hydrophile Stoffe brauchen dafür selektive Proteinkanäle:
- K+ - Kanäle
- Na+ - Kanäle
- Cl- - Kanäle
Der Transport durch Kanäle folgt dem Konzentrationsgradienten und/oder
dem elektrischen Gradienten. Es ist ein passiver Transport, der keine Energie
benötigt. Wir unterscheiden zwei Arten von Transport:
Antiport
Ein Ion geht in die Zelle hinein, ein anderes gleichzeitig auf dem gleichen
Weg entgegengesetzt hinaus.
Cotransport
Transport mehrere Ionen in die gleiche Richtung z.B. Na+
/K+ /2Cl-. Ein wichtiger Cotransport
für Niere und Darm ist der Na+ /Glukose-Cotransport.
Er ist spannungsabhängig weil nur + transportiert wird.
Der Natriumgradient hilft dabei, die Glukose in die Zelle zu befördern.
Spezifische Substanzen erlauben, bestimmte Kanäle zu blockieren. Der
Antiport Cl-/HCO3-; wird z.B. durch SITS blockiert.
Der Cotransport Na+ /K+ /2Cl-
kann durch die Diuretika Furosemid oder Bumetanid blockiert werden.
Nicht erregbare Zellen besitzen viel mehr K+ - Kanäle
als Na+ - Kanäle. Die Zellmembranen sind deshalb
sehr permeabel für K+ und wenig permeabel für
Na+ . Die Permeabilität von Cl-/HCO3-
ist gewöhnlich recht gross, ungefähr in der Grössenordnung
der Permeabilität von K+ .
Man beobachtet, dass wenn die Energiequelle der Zellen blockiert werden,
die Zellen grösser werden und schliesslich platzen. Na+
diffundiert in diesem Falle in die Zelle (wegen Konzentrationsunterschied),
aus Elektroneutralitätsgründen begleitet von Cl-, was
einen sekundären Wasserfluss zur Folge hat ›
Zytolyse.
Aktiver Transport
Ein Transport, der gegen den elektrochemischen Gradienten erfolgt, bezeichnet
man als aktiven Transport. Er verbraucht im Gegensatz zur Diffusion Energie.
Diese wird in Form von ATP bereitgestellt. Der aktive Transport von Na+
von innen nach aussen wird durch die Na+ / K+ - Pumpe
bewerkstelligt. Der Natrium-Ausstrom ist mit dem Kalium-Einstrom gekoppelt.
Die Pumpe erlaubt, dass 3 Na+ die Zelle verlassen und gleichzeitig
2 K+ in die Zelle gelangen, entfernt also in jedem Pumpzyklus
eine Ladung aus der Zelle. Sie ist somit stromerzeugend. Sie stellt damit
intrazellulär die niedrige Na+ - und die hohe K+
-Konzentration sicher. Enzym des Pump-mechanismuses ist die Na-K-ATPase.
Die Na+ / K+ - Pumpe kann durch das Herzglykosid Ouabain
gehemmt werden › Zellen platzen
In einer Zelle befinden sich noch andere Pumpen:

Allgemein wird der aktive Transport über Pumpen gehemmt oder sogar
unterbrochen, wenn die Zelle keine Energie zur Verfügung hat. Das trifft
vorallem in folgenden Fällen zu:

Membranpotential
Unter Membranpotential versteht man eine elektrische Potentialdifferenz
zwischen dem Innern der Zelle und der extrazellulären Flüssigkeit.
Bei einer Zelle misst man ein Membranpotential von 70 - 90 mV. Das
Innere der Zelle ist gegenüber dem Aeusseren negativ gelanden. Auf
der Innenseite der Membran herrscht ein Anionenüberschuss. Extrazellulär
herrscht ein Kationenüberschuss und demnach ist es dort positiv. Die
Zellmembran ist sehr permeabel für K+ und Cl- wenig permeabel für
Na+ und undurchlässig für Proteine und organische Phosphate.

Permeabilität für K+ › Permeabilität
für Na+
Für die Na+/K+ - Pumpe wird Energie gebraucht.
Die Zelle befindet sich folglich nicht in einem Gleichgewichtszustand (normal
braucht Gleichgewicht keine Energie!), sondern in einem stationären
Zustand, dem sogenannten steady state, d.h. in einem durch Energieverbrauch
aufrechterhaltenen Ungleichgewicht. Die Potentialdifferenz zwischen Innen
und Aussen ist folglich kein Nernst-Gleichgewicht, sondern ein Donnan-Gleichgewicht.
Welches Ion bestimmt das Membranpotential ?
Wir haben gesehen, dass falls ein Ion viel besser diffundiert (K+
und Cl-) als ein anderes (Na+) , sich die Gleichung
von Goldmann zur Gleichung von Nernst verkürzt. Danach können
folgende Gleichgewichtspotentiale (E) ausgerechnet werden (siehe Gleichung
4). Hierbei ist der Einstrom gleich dem Ausstrom. Der Nettofluss ist also
gleich null.
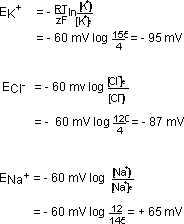
Daraus wir ersichtlich, dass das Membranpotential sehr nahe am Gleichgewichtspotential
von K+ und Cl- ist. Der Einfluss von Cl- ist jedoch eher gering. K+ bestimmt
das Membranpotential.
Zusammenfassend ergibt sich das steady state aus:
- grossem Konzentratinsgradienten für K+
- einer Zellmembran, die für K+ sehr permeabel ist ›
K+ versucht, die Zelle zu verlassen
- einem Zellinnern, das elektrisch negativ geladen ist
- einem elektrochemischen Gradienten, welcher den Einfluss von Na+
in die Zelle begünstigt; trotzdem bleibt die Permeabilität für
Na+ nur gering
- dem wenigen Na+, das in die Zelle eintritt, welches aktiv
über die Na+/ K+ - Pumpe wieder nach aussen
befördert wird (2 K+ hinein für 3 Na+
hinaus)
- dem durch die Na+/ K+-Pumpe generierten Strom,
der für das Membranpotential mitverantwortlich ist.
Elektrisches Membranmodell
Die lipiden Anteile der Membran verleiht ihr die Eigenschaft eines elektrischen
Isolators mit einer Kapazität (C). Die Proteinkanäle lassen mehr
oder weniger einfach Ionen durch. Sie werden deshalb als elektrische Widerstände
(R) angesehen. Die Kanäle stehen in Serie zueinander. Die Leitfähigkeit
(1/R) ist ein Indiz für die Membranpermeabilität.
Insgesammt gibt es 4 Ströme:
- Kapazitiver Strom durch den Kondensator (fettschicht der Zellmembran)
- Stromfluss durch den K+ - Kanal
- Stromfluss durch den Na+ - Kanal
- Stromfluss durch den Cl- - Kanal
Hormone
Hormone dienen, im Gegensatz zu den Nerven, der langsamen internen Kommunikation.
Mit den Hormonen kann die Funktion der Organe angepasst werden. Sie werden
von endokrinen Drüsen produziert, sekretiert und über die Blutbahn
zur extrazellulären Flüssigkeit transportiert. Jedes Hormon reagiert
mit genau bestimmten Zellen (Zielzellen) in einer spezifischen Art. Die
Hormonsekretion wird über eine negative Rückkoppelung geregelt
(z.B. steigt im Falle eines Blutzuckeranstiegs sogleich auch die Sekretion
von Insulin ins Blut). Viele Hormone werden von der Hypophyse freigesetzt.
Regulierung geschieht im Hypothalamus, d.h. wieviele Hormone in der Hypophyse
freigesetzt werden.
Zwei Arten von Hormone werden unterschieden:
Steroidhormone und Schilddrüsenhormone
Steroid- und Schilddrüsenhormone sind fettlöslich und permeabel
im Gegensatz zu anderen Hormongruppen. Sie treten in alle Zellen ein, auch
in diejeningen, in denen sie keine Wirkung haben. Ihre Rezeptoren befinden
sich im Zellinnern, d.h. im Zytoplasma. Diese Hormone wirken auf die Proteinsynthese;
das bedeutet also eine eher langsame Reaktion. Wirkung tritt erst nach Stunden
auf (Latenzzeit ist gross).
Beispiele:
- Sexualhormone: Oestrogen bei Frauen und Testosteron bei Männer
- Schilddrüsenhormone: Thyroxin
- Nebennierenhormone: - Kortikosteroide gebildet in Nebennierenrinde
- Aldosteron, wichtiges Hormon der Niere; wirkt auf Na+-Pumpe und Permeabilität
von Na+
Peptidhormone
Peptidhormone z.B. Hypophysenhormone, Insulin, Glukagon, Calcitonin, Parathormon,
etc. sind nicht fettlöslich. Sie können nicht einfach durch die
Zellmembran diffundieren. Ihre Rezeptoren sitzen auf der Zellmembran.
Genereller Reaktionsmechanismus
- Biosynthese, Lagerung und Sekretion durch endokrine Drüsen
- negative Rückkoppelung und zugleich Abbau des Hormons durch die
Zelle
- die Zahl der Rezeptoren ist begrenzt, die Hormonkonzentration ist variabel

Ist K gross, bindet sich das Hormon gut mit dem Rezeptor.
Die Effizienz E eines Hormons ist abhängig von seiner Affinität
zum Rezeptor, von der Konzentration des Hormons und von der Konzentration
des Rezeptors.
[HR] = K · [H] · [R].
Gewisse Substanzen können sich, ohne eine Wirkung auszulösen,
am gleichen Rezeptor binden. Diese nennt man kompetitive Inhibitoren.
Der Effekt eines Hormons kann durch die Wirkung eines anderen Hormons wieder
aufgehoben werden (nicht am gleichen Rezeptor anheftend) = nicht-kompetitive
Inhibitoren.
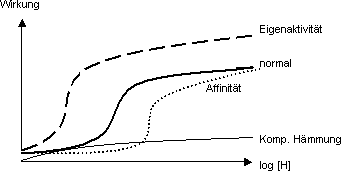
Eigenaktivität: max. Hormoneffekt
Wirksamkeit des Hormons unter Bedingung, dass Rezeptor an Hormon gebunden
ist
Kompetitive Hämmung: weil Eigenaktivität = 0 ist Kurve flach
Nicht-kompetitive Hämmung: nicht auf Niveau Rezeptoren, sondern auf
Niveau der Zelle; sowohl Hormon wie komp. Hämmung kann sich mit Rezeptor
binden
Affinität Kurve verschiebt sich gegen grössere Konzentrationen:
es braucht mehr, um dass wir dieselben Hormon-Rezeptor- Komplexe bekommen
› für gleichen Hormoneffekt braucht es grössere
Konzentrationen
Reaktionsablauf bei Steroid- und Schilddrüsenhormonen
Da die Steroid- und Schilddrüsenhormone fettlöslich sind, diffundieren
sie ungehindert durch die Zelle. Sie lösen einen Reiz im Zytoplasma
oder Zellkern aus. Der Rezeptor kann sich nach der Hormonbindung z.B. an
das Chromatin binden › Modifikation der Transkription der mRNA ›
spezielle Actine werden produziert, welche ihrerseits wiederum die Synthese
von spezifischen intrazellulären Proteine anregen › diese Enzyme
oder Membranproteine manifestieren die Wirkung der Hormone.
So haben die Sexualhormone und das Kortison eine vielfältige Wirkung
auf den Zellmetabolismus in den verschiedenen Geweben.
Die Latenzzeit von Steroidhormonen beträgt ca. 2-6 Stunden, wobei
vorallem die Proteinsynthese am meisten Zeit konsumiert. Die Zeit bis zur
Anlagerung des Hormons an seinen Rezeptor im Zytosol oder Zellkern beträgt
nur ca. 5 Minuten.
Der Hormoneffekt kann durch Substanzen, welche sich an den Rezeptoren binden
gehemmt werden. Klomifen z.B. bindet sich an Oestrogenrezeptoren (Behandlung
von Sterilität und Brustkrebs). Spironolacton bindet sich an den Aldosteronrezeptor
(wird als Diuretikum eingesetzt).
Reaktionsablauf bei Peptidhormonen
Da Peptidhormone die Zellmembran nicht durchqueren können, wird ihr
Effekt duch ein Inter-medium, d.h. „eine spezifische Rezeptorbindung„
hervorgerufen. Diese Bindung aktiviert ein spezifisches Membranprotein mit
dem Namen G-Protein. Dieses wiederum aktiviert oder hemmt weitere Membranproteine.
Diese Reaktionen führen im Allgemeinen zur Bildung eines second Messengers,
der intrazellulär wirkt. Der erste Messenger ist das Hormon selbst.
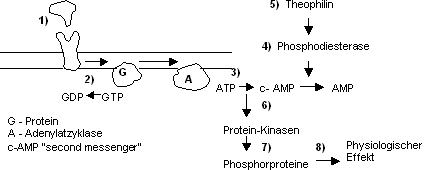
Schematisch sieht die Reaktion des Peptidhormons wie folgt aus:
- Bindung des Hormons am Rezeptor auf der Membran
- Aktivierung des membranösen G-Proteins durch Bindung von Guanosintriphosphat
(GTP), GDP wird frei. Lässt die Hormonwirkung wieder nach, wird GTP
wieder frei.
- Konfiguration des G-Proteins ändert, hat somit Wirkung auf die
das Enzym Adenylatzyklase. Dieses bildet aus ATP den zweiten Messenger,
cyclo-AMP,
- welcher duch das Enzym Phosphodiesterase aufgelöst wird.
- Durch Zugabe von Theophilin oder verwandte Stoffe, wie Coffein kann
die Phosphodiesterase blockiert werden, c-AMP kann nicht mehr abgebaut
werden. C-AMP häuft sich an › Hormoneffekt hält an. Theophilin
als Verstärker des Hormoneffektes.
- C-AMP aktiviert Proteinkinasen, die auf den zellulären Metabolismus
Einfluss nehmen .
- Diese aktivieren ihrerseits Phosphorproteine, die z.B. Membrankanäle
öffnen können ›
- physiologischer Effekt durch Ionenströme
Die Latenzzeit zwischen dem Ausschütten des Peptidhormons und seinem
bemerkbaren Effekt beträgt, im Gegensatz zu den Steroidhomonen, nur
gerade einige Minuten.
Kompetitive Hemmung
Bei der nichtkompetitiven Hemmung besitzt das hemmende Hormon einen eigenen
Rezeptor. Nach der Andockung des Hormons am Rezeptor kommt es zur Bindung
von GTP im G-Protein. Dies inaktiviert die Adenylatzyklase, wodurch es zu
einer Hemmung der c-AMP-Bildung kommt. Der hormonelle Effekt bleibt aus.
Verschiedene Peptidhormone können gegensätzliche Effekte haben.
Während das Peptidhormon Insulin den Blutzuckergehalt senkt, bewirkt
das Glukagon das Gegenteil. Das Parathormon erhöht die Konzentration
von Kalzium im Blut, während Calzitonin die gegenteilige Wirkung zeigt.
Im Moment sind keine pharmakologischen Substanzen im Umlauf, welche als
kompetitive Inhibitoren für Peptidhormone gebraucht werden. Bei den
Catecholaminen ( Adrenalin), welche die gleichen Wirkungsmechanismen haben
wie die Peptidhormone, gibt es zahlreiche Substanzen mit kompetitiver Wirkung.
|
